
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 157
Title
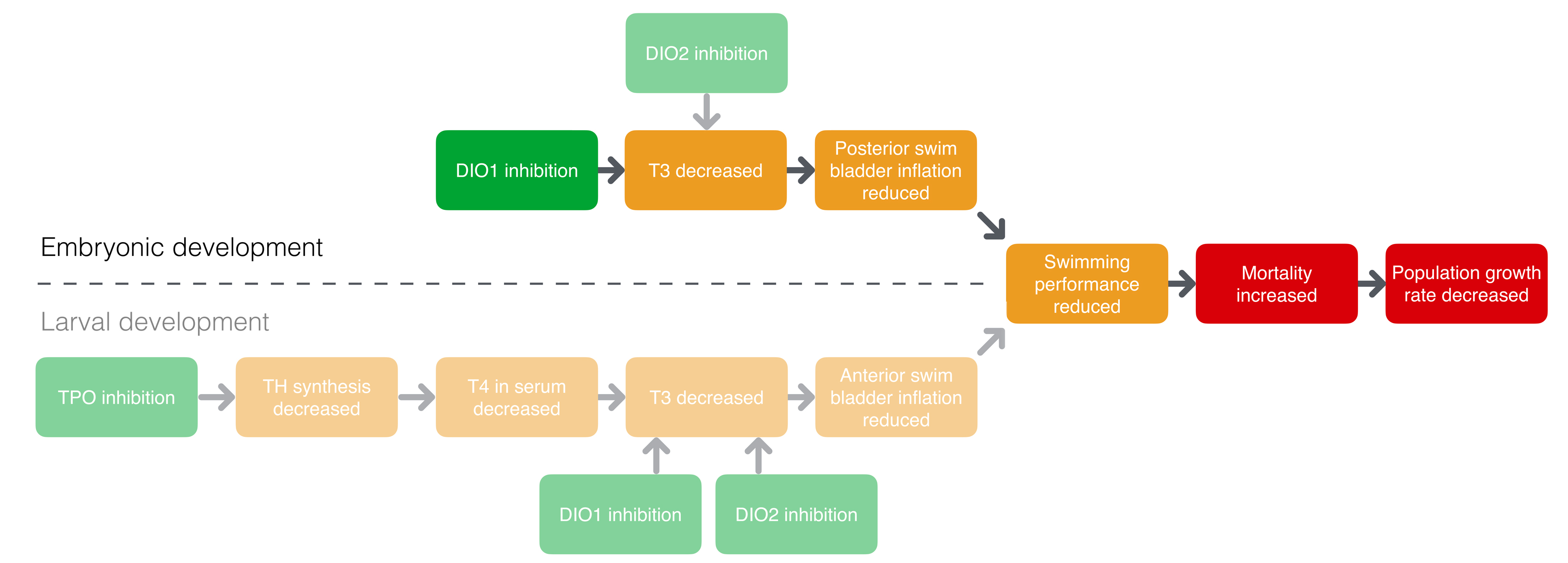
Deiodinase 1 inhibition leading to increased mortality via reduced posterior swim bladder inflation
Short name
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 157, Deiodinase 1 inhibition leading to increased mortality via reduced posterior swim bladder inflation in tools offered by third parties.
Point of Contact
Contributors
- Dries Knapen
- Lucia Vergauwen
- Ann-Cathrin Haigis
- Agnes Aggy
Coaches
- Shihori Tanabe
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
| 1.35 | WPHA/WNT Endorsed |
This AOP was last modified on May 26, 2024 20:39
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Inhibition, Deiodinase 1 | October 10, 2022 08:07 |
| Decrease, Population growth rate | January 03, 2023 09:09 |
| Decreased, Triiodothyronine (T3) | October 07, 2022 08:26 |
| Reduced, Posterior swim bladder inflation | October 07, 2022 08:42 |
| Reduced, Swimming performance | September 08, 2021 06:12 |
| Increased Mortality | July 08, 2022 07:32 |
| Inhibition, Deiodinase 1 leads to Decreased, Triiodothyronine (T3) | October 10, 2022 08:15 |
| Decreased, Triiodothyronine (T3) leads to Reduced, Posterior swim bladder inflation | October 07, 2022 09:12 |
| Reduced, Posterior swim bladder inflation leads to Reduced, Swimming performance | September 03, 2021 13:02 |
| Reduced, Swimming performance leads to Increased Mortality | September 08, 2021 10:05 |
| Increased Mortality leads to Decrease, Population growth rate | July 08, 2022 08:29 |
| Inhibition, Deiodinase 1 leads to Reduced, Posterior swim bladder inflation | October 10, 2022 08:23 |
| Reduced, Posterior swim bladder inflation leads to Increased Mortality | September 08, 2021 10:08 |
Abstract
This AOP describes the sequence of events leading from deiodinase inhibition to increased mortality via reduced posterior swim bladder inflation. Thyroid hormones (THs) are critical during embryonic development and disruption of the TH system can interfere with normal development. Three types of iodothyronine deiodinases (DIO1-3) have been described in vertebrates that activate or inactivate THs and are therefore important mediators of TH action. While type II deiodinase (DIO2) has thyroxine (T4) as a preferred substrate and is mostly important for converting T4 to the more biologically active triiodothyronine (T3), type I deiodinase is capable of both converting T4 into T3 and converting rT3 to 3,3’ T2. Inhibition of DIO1 thus reduces T3 levels. However, partly because rT3, rather than T4, is the preferred substrate for DIO1, DIO1 inhibition is probably less important in causing reduced T3 levels when compared to DIO2 inhibition. As in amphibians, the transition between the different developmental phases in fish, including maturation and inflation of the swim bladder, is mediated by THs (Brown et al., 1988; Liu and Chan, 2002). The swim bladder is a gas-filled organ that typically consists of two chambers (Robertson et al., 2007). The posterior chamber inflates during early development in the embryonic phase, while the anterior chamber inflates during late development in the larval phase. This AOP describes how DIO1 inhibition results in reduced T3 levels, which prohibit normal inflation of the posterior chamber of the swim bladder in the embryonic phase. The posterior chamber is important for regulating buoyancy and thus for swimming performance (Robertson et al., 2007). Reduced swimming performance reduces chances of survival due to a decreased ability to forage and avoid predators. The final adverse outcome is a decrease of the population trajectory. Since many AOPs eventually lead to this more general adverse outcome at the population level, the more specific and informative adverse outcome at the organismal level, increased mortality, is used in the AOP title. Support for this AOP is mainly based on chemical exposures in zebrafish and fathead minnows (Jomaa et al., 2014; Cavallin et al., 2017; Stinckens et al., 2018) and on knockdown studies of combined inactivation of DIO1 and DIO2 in zebrafish embryos (Walpita et al., 2009, 2010; Heijlen et al., 2014; Bagci et al., 2015).
This AOP is part of a larger AOP network describing how decreased synthesis and/or decreased biological activation of THs leads to incomplete or improper inflation of the swim bladder, leading to reduced swimming performance, increased mortality and decreased population trajectory (Knapen et al., 2018; Knapen et al., 2020; Villeneuve et al., 2018). Other than the difference in deiodinase (DIO) isoform, the current AOP is identical to the corresponding AOP leading from DIO2 inhibition to increased mortality via posterior swim bladder inflation (https://aopwiki.org/aops/155). The overall importance of DIO1 versus DIO2 in fish is not exactly clear. DIO1 inhibitors are often also inhibitors of DIO2 (Olker et al., 2019; Stinckens et al. 2018). In the ToxCast DIO1 inhibition single concentration assay, 219 out of 1820 chemicals were positive and 177 of these were also positive for DIO2 inhibition (viewed on 5/7/2022). This complicates the distinction between the relative contribution of DIO1 and DIO2 inhibition to reduced swim bladder inflation. The current state of the art suggests that DIO2 is more important than DIO1 in regulating swim bladder inflation. Therefore AOP 155 may be of higher biological relevance compared to the AOP that is described here
AOP Development Strategy
Context
The larger AOP network describing the effect of deiodinase and thyroperoxidase inhibition on swim bladder inflation consists of 5 AOPs:
- Deiodinase 2 inhibition leading to increased mortality via reduced posterior swim bladder inflation: https://aopwiki.org/aops/155
- Deiodinase 2 inhibition leading to increased mortality via reduced anterior swim bladder inflation: https://aopwiki.org/aops/156
- Deiodinase 1 inhibition leading to increased mortality via reduced posterior swim bladder inflation : https://aopwiki.org/aops/157
- Deiodinase 1 inhibition leading to increased mortality via reduced anterior swim bladder inflation : https://aopwiki.org/aops/158
- Thyroperoxidase inhibition leading to increased mortality via reduced anterior swim bladder inflation: https://aopwiki.org/aops/159
The development of these AOPs was mainly based on a series of dedicated experiments (using a set of reference chemicals as prototypical stressors) in zebrafish and fathead minnow that form the core of the empirical evidence. Specific literature searches were used to add evidence from other studies, mainly in zebrafish and fathead minnow. No systematic review approach was applied.
Strategy
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 1009 | Inhibition, Deiodinase 1 | Inhibition, Deiodinase 1 |
| KE | 1003 | Decreased, Triiodothyronine (T3) | Decreased, Triiodothyronine (T3) |
| KE | 1004 | Reduced, Posterior swim bladder inflation | Reduced, Posterior swim bladder inflation |
| KE | 1005 | Reduced, Swimming performance | Reduced, Swimming performance |
| AO | 351 | Increased Mortality | Increased Mortality |
| AO | 360 | Decrease, Population growth rate | Decrease, Population growth rate |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Inhibition, Deiodinase 1 leads to Decreased, Triiodothyronine (T3) | adjacent | Low | Low |
| Decreased, Triiodothyronine (T3) leads to Reduced, Posterior swim bladder inflation | adjacent | Moderate | Low |
| Reduced, Posterior swim bladder inflation leads to Reduced, Swimming performance | adjacent | Moderate | Low |
| Reduced, Swimming performance leads to Increased Mortality | adjacent | Moderate | Low |
| Increased Mortality leads to Decrease, Population growth rate | adjacent | Moderate | Moderate |
| Inhibition, Deiodinase 1 leads to Reduced, Posterior swim bladder inflation | non-adjacent | Moderate | Low |
| Reduced, Posterior swim bladder inflation leads to Increased Mortality | non-adjacent | High | Low |
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Embryo | High |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Overall Assessment of the AOP

The attached document includes:
- Support for biological plausibility of KERs
- Support for essentiality of KEs
- Empirical support for KERs
- Dose and temporal concordance table covering the larger AOP network
Overall, the weight of evidence for the sequence of key events laid out in the AOP is moderate, and it should be noted that based on available evidence DIO2 seems to be more important than DIO1 in providing sufficient T3 for swim bladder inflation. The exact underlying mechanism of TH disruption leading to impaired swim bladder inflation is not exactly understood.
Domain of Applicability
Life stage: The current AOP is only applicable to early embryonic development, which is the period where the posterior swim bladder chamber inflates. In all life stages, the conversion of T4 into more biologically active T3 by DIO1 is essential. Inhibition of DIO1 therefore impacts swim bladder inflation in both early and late (https://aopwiki.org/aops/158) developmental life stages.
Taxonomic: Organogenesis of the swim bladder begins with an evagination from the gut. In physostomous fish, a connection between the swim bladder and the gut is retained. In physoclystous fish, once initial inflation by gulping atmospheric air at the water surface has occurred, the swim bladder is closed off from the digestive tract and swim bladder volume is regulated by gas secretion into the swim bladder (Woolley and Qin, 2010). This AOP is currently mainly based on experimental evidence from studies on zebrafish and fathead minnows, physostomous fish with a two-chambered swim bladder. Knowledge could be expanded to physoclistous fish, such as the Japanese rice fish or medaka (Oryzias latipes) that has a single chambered swim bladder that inflates during early development.
Sex: All key events in this AOP are plausibly applicable to both sexes. Sex differences are not often investigated in tests using early life stages of fish. In medaka, sex can be morphologically distinguished as soon as 10 days post fertilization. Females appear more susceptible to thyroid‐induced swim bladder dysfunction compared with males (Godfrey et al., 2019). In zebrafish and fathead minnow, it is currently unclear whether sex-related differences are important in determining the magnitude of the changes of the sequence of events along this AOP. Different fish species have different sex determination and differentiation strategies. Zebrafish do not have identifiable heteromorphic sex chromosomes and sex is determined by multiple genes and influenced by the environment (Nagabhushana and Mishra, 2016). Zebrafish are undifferentiated gonochorists since both sexes initially develop an immature ovary (Maack and Segner, 2003). Immature ovary development progresses until approximately the onset of the third week. Later, in female fish immature ovaries continue to develop further, while male fish undergo transformation of ovaries into testes. Final transformation into testes varies among male individuals, however finishes usually around 6 weeks post fertilization. Since the posterior chamber inflates around 5 days post fertilization in zebrafish, when sex differentiation has not started yet, sex differences are expected to play a minor role in the current AOP. Fathead minnow gonad differentiation also occurs during larval development. Fathead minnows utilize a XY sex determination strategy and markers can be used to genotype sex in life stages where the sex is not yet clearly defined morphologically (Olmstead et al., 2011). Ovarian differentiation starts at 10 dph followed by rapid development (Van Aerle et al., 2004). At 25 dph germ cells of all stages up to the primary oocytes stage were present and at 120 dph, vitellogenic oocytes were present. The germ cells (spermatogonia) of the developing testes only entered meiosis around 90–120 dph. Mature testes with spermatozoa are present around 150 dph. Since the posterior chamber inflates around 6 days post fertilization (1 dph) in fathead minnows, sex differences are expected to play a minor role in the current AOP.
Essentiality of the Key Events
Overall, the support for essentiality of the KEs is low. There is ample evidence from combined DIO1 and DIO2 knockdown studies in zebrafish that shows downstream effects, and evidence from both chemical exposure with TH supplementation and knockdown with TH supplementation showing that blocking a KE prevents downstream KEs from occurring. There is no specific evidence for the essentiality of DIO1 inhibition independent of DIO2 inhibition and DIO2 seems more important than DIO1 in providing sufficient T3 for proper swim bladder inflation. Therefore the overall evidence for essentiality is considered low.
Evidence Assessment
Biological plausibility: see Table. Overall, the weight of evidence for the biological plausibility of the KERs in the AOP is moderate since there is empirical support for an association between the sets of KEs and the KERs are plausible based on analogy to accepted biological relationships, but scientific understanding is not completely established.
Empirical support: see Table. Overall, the empirical support for the KERs in the AOP is moderate since dependent changes in sets of KEs following exposure to several specific stressors has been demonstrated, with limited evidence for dose and temporal concordance and some uncertainties.
Known Modulating Factors
Quantitative Understanding
Quantitative understanding of this AOP is currently lacking.
Considerations for Potential Applications of the AOP (optional)
A growing number of environmental pollutants are known to adversely affect the thyroid hormone system, and major gaps have been identified in the tools available for the identification, and the hazard and risk assessment of these thyroid hormone disrupting chemicals. Villeneuve et al. (2014) discussed the relevance of swim bladder inflation as a potential key event and endpoint of interest in fish tests. Knapen et al. (2020) provide an example of how the adverse outcome pathway (AOP) framework and associated data generation can address current testing challenges in the context of fish early-life stage tests, and fish tests in general. While the AOP is only applicable to fish, some of the upstream KEs are relevant across vertebrates. The taxonomic domain of applicability call of the KEs can be found on the respective pages. A suite of assays covering all the essential biological processes involved in the underlying toxicological pathways can be implemented in a tiered screening and testing approach for thyroid hormone disruption in fish, using the levels of assessment of the OECD’s Conceptual Framework for the Testing and Assessment of Endocrine Disrupting Chemicals as a guide. Specifically, for this AOP, deiodinase inhibition can be assessed using an in chemico assay, measurements of T3 levels could be added to the Fish Embryo Acute Toxicity (FET) test (OECD TG 236), the Fish Early Life Stage Toxicity (FELS) Test (OECD TG210) and the Fish Sexual Development test (FSDT) (OECD TG 234), and assessments of posterior chamber inflation and swimming performance could be added to the FELS Test and FSDT.
Thyroid hormone system disruption causes multiple unspecific effects. Addition of TH measurements could aid in increasing the diagnostic capacity of a battery of endpoints since they are specific to the TH system. A battery of endpoints would ideally include the MIE, the AO and TH levels as the causal link. It is also in this philosophy that TH measurements are currently being considered as one of the endpoints in project 2.64 of the OECD TG work plan, “Inclusion of thyroid endpoints in OECD fish Test Guidelines”. While T3 measurements showed low levels of variation and were highly predictive of downstream effects in dedicated experiments to support this AOP, more variability may be present in other studies. Because of the rapid development in fish, it is important to compare T3 levels within specific developmental stages. For example, clear changes in T3 levels have been observed in zebrafish at 14, 21 and 32 dpf (Stinckens et al., 2020) and in fathead minnows at 4, 6, 10, 14, 18 and 21 dpf (Nelson et al., 2016; Cavallin et al., 2017) using liquid chromatography tandem mass spectrometry (LC−MS/MS).
The overall importance of DIO1 versus DIO2 in fish is not exactly clear. The current state of the art suggests that DIO2 is more important than DIO1 in regulating swim bladder inflation. Therefore AOP 155 may be more relevant for applications compared to the AOP that is described here.
References
Bagci, E., Heijlen, M., Vergauwen, L., Hagenaars, A., Houbrechts, A.M., Esguerra, C.V., Blust, R., Darras, V.M., Knapen, D., 2015. Deiodinase knockdown during early zebrafish development affects growth, development, energy metabolism, motility and phototransduction. PLOS One 10, e0123285.
Brown, C.L., Doroshov, S.I., Nunez, J.M., Hadley, C., Vaneenennaam, J., Nishioka, R.S., Bern, H.A., 1988. MATERNAL TRIIODOTHYRONINE INJECTIONS CAUSE INCREASES IN SWIMBLADDER INFLATION AND SURVIVAL RATES IN LARVAL STRIPED BASS, MORONE-SAXATILIS. Journal of Experimental Zoology 248, 168-176.
Cavallin, J.E., Ankley, G.T., Blackwell, B.R., Blanksma, C.A., Fay, K.A., Jensen, K.M., Kahl, M.D., Knapen, D., Kosian, P.A., Poole, S.T., Randolph, E.C., Schroeder, A.L., Vergauwen, L., Villeneuve, D.L., 2017. Impaired swim bladder inflation in early life stage fathead minnows exposed to a deiodinase inhibitor, iopanoic acid. Environmental Toxicology and Chemistry 36, 2942-2952.
Godfrey A, Hooser B, Abdelmoneim A, Sepulveda MS. 2019. Sex-specific endocrine-disrupting effects of three halogenated chemicals in japanese medaka. Journal of Applied Toxicology. 39(8):1215-1223.
Heijlen, M., Houbrechts, A., Bagci, E., Van Herck, S., Kersseboom, S., Esguerra, C., Blust, R., Visser, T., Knapen, D., Darras, V., 2014. Knockdown of type 3 iodothyronine deiodinase severely perturbs both embryonic and early larval development in zebrafish. Endocrinology 155, 1547-1559.
Jomaa, B., Hermsen, S.A.B., Kessels, M.Y., van den Berg, J.H.J., Peijnenburg, A.A.C.M., Aarts, J.M.M.J.G., Piersma, A.H., Rietjens, I.M.C.M., 2014. Developmental Toxicity of Thyroid-Active Compounds in a Zebrafish Embryotoxicity Test. Altex-Alternatives to Animal Experimentation 31, 303-317.
Knapen, D., Angrish, M.M., Fortin, M.C., Katsiadaki, I., Leonard, M., Margiotta-Casaluci, L., Munn, S., O'Brien, J.M., Pollesch, N., Smith, L.C., Zhang, X.W., Villeneuve, D.L., 2018. Adverse outcome pathway networks I: Development and applications. Environmental Toxicology and Chemistry 37, 1723-1733.
Knapen, D., Stinckens, E., Cavallin, J.E., Ankley, G.T., Holbech, H., Villeneuve, D.L., Vergauwen, L., 2020. Toward an AOP Network-Based Tiered Testing Strategy for the Assessment of Thyroid Hormone Disruption. Environmental Science & Technology 54, 8491-8499.
Liu, Y.W., Chan, W.K., 2002. Thyroid hormones are important for embryonic to larval transitory phase in zebrafish. Differentiation 70, 36-45.
Maack, G., Segner, H., 2003. Morphological development of the gonads in zebrafish. Journal of Fish Biology 62, 895-906.
Nagabhushana A, Mishra RK. 2016. Finding clues to the riddle of sex determination in zebrafish. Journal of Biosciences. 41(1):145-155.
Olmstead AW, Villeneuve DL, Ankley GT, Cavallin JE, Lindberg-Livingston A, Wehmas LC, Degitz SJ. 2011. A method for the determination of genetic sex in the fathead minnow, pimephales promelas, to support testing of endocrine-active chemicals. Environmental Science & Technology. 45(7):3090-3095.
Robertson, G.N., McGee, C.A.S., Dumbarton, T.C., Croll, R.P., Smith, F.M., 2007. Development of the swimbladder and its innervation in the zebrafish, Danio rerio. Journal of Morphology 268, 967-985.
Stinckens, E., Vergauwen, L., Ankley, G.T., Blust, R., Darras, V.M., Villeneuve, D.L., Witters, H., Volz, D.C., Knapen, D., 2018. An AOP-based alternative testing strategy to predict the impact of thyroid hormone disruption on swim bladder inflation in zebrafish. Aquatic Toxicology 200, 1-12.
van Aerle R, Runnalls TJ, Tyler CR. 2004. Ontogeny of gonadal sex development relative to growth in fathead minnow. Journal of Fish Biology. 64(2):355-369.
Villeneuve, D., Angrish, M., Fortin, M., Katsiadaki, I., Leonard, M., Margiotta-Casaluci, L., Munn, S., O'Brien, J., Pollesch, N., Smith, L., Zhang, X., Knapen, D., 2018. Adverse Outcome Pathway Networks II: Network Analytics. Environ Toxicol Chem doi: 10.1002/etc.4124.
Villeneuve, D., Volz, D.C., Embry, M.R., Ankley, G.T., Belanger, S.E., Leonard, M., Schirmer, K., Tanguay, R., Truong, L., Wehmas, L., 2014. Investigating alternatives to the fish early-life stage test: a strategy for discovering and annotating adverse outcome pathways for early fish development. Environmental Toxicology and Chemistry 33, 158-169.
Walpita, C.N., Crawford, A.D., Darras, V.M., 2010. Combined antisense knockdown of type 1 and type 2 iodothyronine deiodinases disrupts embryonic development in zebrafish (Danio rerio). Gen Comp Endocrinol 166, 134-141.
Walpita, C.N., Crawford, A.D., Janssens, E.D., Van der Geyten, S., Darras, V.M., 2009. Type 2 iodothyronine deiodinase is essential for thyroid hormone-dependent embryonic development and pigmentation in zebrafish. Endocrinology 150, 530-539.
Woolley, L.D., Qin, J.G., 2010. Swimbladder inflation and its implication to the culture of marine finfish larvae. Reviews in Aquaculture 2, 181-190.